El cacao es uno de los cultivos que esta intimamente ligado a Ia tradicion y Ia cultura Mexicana y es uno de los productos domesticados por los antiguos habitantes de Mexico. Las técnicas de propagacion son importantes en este cultivo ya que de estas depende el éxito 0 el fracaso de Ia productividad de las plantaciones. El mejoramiento de Ia propagaciOn vegetativa de esta especie es de gran mmportancma debido a que las tecnicas horticolas actualmente disponibles no alcanzan a cubrir Ia demanda de plantas requeridas para Ia renovaciOn de las plantaciones establecidas. Dado el potencial que representa Ia embriogenesis somatica para a clonacion del cacao, en diversos centros de investigacmon del mundo Se desarrollan expermencias tendientes a su perfeccionamiento. En esta comunicacion Se presentan resultados de experiencias desarrolladas con Ia finalidad de adaptar on protolo previamente desarrollado a a clonacion de genotipos seleccionados en Mexico. Los factores estudmados fueron el efecto de reguladores del crecimiento auxinas y citoquininas, azUcares, sales minerales y pretratamientos a bajas temperaturas de explantes antes de a siembra. Como explantes Se utilizaron piezas florales (pétalos, estaminodios, filamentos de anteras) que fueron tratados y cultivados de acuerdo a on protocolo desarrollado previamente. Los mejores resultados de induccion de tejido embriogenico Se obtuvieron en explantes cultivados en S medio nutritivo “MS’ modificado, enriquecido con glucosa 50g11 y ajustandose a on pH de 5,5. La mejor composicion hormonal favorable a Ia induccion en el medio de indoccion primario resulto de las combinaciones de 2,4,5-T (1mg/I) o 2,4-D (1mg/I) + kmnetina (0,25mg11) durante 3 semanas. En el medio secundario para Ia expresiOn de a embriogénesis se determmnO que Ia presencia de reguladores de crecimmento no es necesaria para a obtencmon de tejido embriogenico. Los pretratam~entos de bajas temperaturas a los botones florales antes de Ia siembra que resultaron favorables al proceso de embriogenesis fueron una mncubacion durante IS a 20 mm a una temperatora de 0 °C y de 30 y 120 mm a una temperatura de 5 °C. El protocolo modificado foe aplicado al cuitivo de explantes de 12 genotipos Mexicanos, algunos de ellos considerados recalcitrantes, los resultados obtenidos fueron positivos ya que se logro Ia inducciOn de tejido embriogénico en todos los genotipos evaluados, con una tasa de embriogénesis que variO de 1,09% al 41%. De estas experiencias Se concluye que a capacidad embriogénica es el resultado de a interaccion del genotipo del explante, del balance hormonal y de Ia composiciOn mineral y energetica del medio de cultivo en el medio primario de nducción de Ia embriogénesis.
Advances in Cocoa - Theobroma cacao - Propagation by Somatic Embryogenesis in Mexico
Abstract
Cocoa is one of the crops that it is intimately inked to Mexican tradition and culture. It is one of the products that was domesticated by the ancient inhabitants of Mexico. Propagation techniques are important in cocoa cultivation since success or fairure of the plantations is dependent on such techniques. The improvement of micropropagation is of great importance due to the fact that the currently available horticultural techniques do not cover the demand required for the renovation of the established plantations. In this communication, we present results from a series of modifications to a previously developed protocol for regeneration through somatic embryogenesis, and the application of the improved protocol to selected
genotypes in Mexico. Explants were derived from floral parts (petals, staminodes and anther
filaments) that had been treated and cultivated according to the previously developed protocoL The control culture medium, known to give good results for the induction of embryogenic calli, was a modified MS medium enriched with glucose 50 g/l and with the pH adjusted to 5.5. This culture medium was modified to include various carbohydrate sources, minerals and growth hormones to investigate their effect on the induction of embryogenic calli. Glucose at concentrations of 50 and 60g/l and saccharose to Bog/I resulted in the highest frequency of embryogenic caliL Aoxins (2,4,5-T or 2,4-D at 1mg/I) and kinetin (0,25mg/I) also increased the frequency of somatic embryogenic calli. However, it was established that the presence of growth regulators is not necessary for embryogenesis expressmon in the secondary medium. The pretreatment of floral explants before the in vitro culture at temperatures of 0°C for 15 or 20 minutes, or at 5°C for 120 minutes resulted in a increase in the percentage of embryogenic calli formed. These results allowed us to modify the original protocol for embryogenesis and test this modified version on explants from 12 genotypes, some of which are considered as recalcitrant The rates of embryogenesis obtained varied between 1 and 41%. We conclude that embryogenic capacity ms a result of the interaction between the genotype of the explant, the hormonal balance and the mineral and carbohydrate composition of the primary culture medium used to induce embryogenesis.
Introducción
El cacao es on cultivo perenne, alOgamo y tropmcal, del cual en Mexico son cultivadas 90 000 ha distribuidas 60 000 en Tabasco y 30 000 en Chiapas, a prodocción anoal se estina en 43 000 ton do cacao seco. Do este coltivo se benefician de manera directa 35 000 familmas do las quo 12 000 se encuentran en Chiapas y 23 000 en Tabasco, Entre los factores que limitan Ia producciOn del cacao en Mexico destacan:
Corno resultado do investigaciones en el mejoramiento genético, existe material seleccionado con rendimiientos superiores a una ton/ha de cacao seco y de alta resistenceia a la mancha negra; este material constituye la base del programma derenovacion de plantaciones emprendido por el gobiemo mexicano desde 1996. Para la produccion masiva de plantas de clones mejorados de cacao, actualmente se applican las tecnicas de injertado en plnatula y el enraizamiento de ramas, sin embargo, a pesar de la eficiencia de estas tecnicas, no se alcanza a cubrir la demanda de plantas requeridas para la renovacion de las plantaciones, que tan solo para el estado de Chiapas son del orden de cinco milliones de plantas al ano.
En el ambmto agronómico, so reconoce quo el cacao es una especie dificil do propagar el injertado convencional, el enraizado de ramas, el microinjertado en plantula y Ia aplicaciôn de noevas tecnologias como Ia propagación mediante el cultivo de tejidos in vitro (Aguilar et al. 1993; Yow y Lim 1994; Flores y Vera 1995; Lopez 1997),
La embriogénesis sornática in vitro como via do propagacián es el procedimiento que permite Ia obtencion de embriones a partir de tejido somático, es decir, sin Ia intervenciOn do celulas gameticas. Los embriones obtenidos por esta via Ilarnados “somâticos” presentan una morfologia y un desarrollo similares a los embriones obtenidos par Ia fecondacion, pero contrariamente a estos, tienen Ia particularidad de presentar una estructura genética identica a Ia de a planta de origen (Lopez 1996). En el caso del cacao, desde hace ya algonos ańos, se han venido desarrollando investigaciones concernientes a Ia propagaciOn in vitro. La primera obtenciOn embriones somaticos aunque sin Ia obtenciOn de plantas do cacao fue reportada por Esan (1977) a partir tie oxplantes de embriones cigoticos inmaduros; esta aptitud fue posteriormente confirrnada por otros autores (Kononowicz et al 1984; Adu-Ampomah et al 1988; Lizt 1986; Sondahl et al. 1988, Chatelet et al 1992; Pence et at 1989). Con sus nvest~gaciones, demostraron Ia potencialidad de diferentes explantes somAticos como las hojas, las nucelas, el togumento interno del ovario y de los pétalos.
Los resultados obtenidos por Lopez y colaboradores (1993; 1996) mostraron a posibilidad do inducciOn do tejido con aptitud embrmogenica asi como el desarrollo de los embriones somáticos y de plantas aptas al cultivo, a partir del cultivo in vitro do pétalos, estaminodios y filamentos do anteras; una consideracion a tomar en cuenta es quo este protocolo fue desarroliado trabajando con genotipos del grupo genético Forastero. La inducciOn de embriogénesis es efectuada en un medio sOlido complementado con los reguladores del crecimiento 2,4-D y kinetina seguida do una fase tie expresiOn en Un medio sin reguladores. Para so desarrollo, los embriones somâticos son sucesivamente cultivados en medios do maduraciOn, gernhinaciOn y do conversiOn do plantulas, después del cual son transferidos a invernadero para aclimataciOn. La eficiencia do este protocolo ha sido superior tanto en cantidad como en calidad do a embriogénosis, a los reportados anteriormente para cacao.
La conformidad genética ox vitro do las plantas obtenidas por esta via ha sido evaluada por LOpez et at (1996) a través del estudio del nivel de ploidia del ADN nuclear do plantas regeneradas por embriogénesis somática con el protocolo anteriormente senalado; los resultados obtenidos indican que no hobo modifrcaciOn del nivel de ploidia y del contenido de ADN nuclear en el proceso de embriogonesis somática. El hábito do crecimiento y do desarrollo de as plantas obtenidas parece normal y el comportamionto reproductivo do as plantas es similar a las obtenidas tie semilla.
No obstante, a reproducción de este protocolo bajo condiciones do paisos productores de cacao y con genotipos diferentes ha resultado dificil (Alemanno 1995; Lápez eta!, 1997a) por lo quo debido al enorme interés quo representa Ia aplicaciôn a gran escala do esta via a Ia propagación del cacao se ha considerado do vital importancia desarrollar investigaciones para su perfoccionamiento y adecuaciOn a las condiciones y genotipos, como es el caso do Méxmco y de otros centros de investigaciOn.
De lo expuesto anteriormente se deduce que a pesar do quo existen técnicas de clonacmon actualmente disponibles para Ia producciOn rnasiva de plantas de cacao, estas no son suficientes para abastecer Ia demanda; y a nivel mundial se roconoce a necesidad de desarrollar técnicas mâs eficientes para a propagación masiva do ârboles tie cacao de alto valor agronomico e industrial.
Por otra parte, dosde ol punto do vista del mejoramiento genético, a clonacion del cacao so considora Ia prUxima etapa necesaria para optimizar a explotaciOn tie los boneflcmos directos tie un genotipo seleccionado 0 creado para rosponder a necesidades diversas como son Ia resistencia a parasitos, altos rendirnientos, alta calidad chocolatera (Lopez 1996).
En el caso particular de Mexico, existe ol interes por desarrollar a nivel comercial a propagaciôn in vitro, ya que do acuerdo a Ia experiencia do otros cultivos, esta presenta un potencial mayor para Ia multiplicacion do genotipos seloccionados, Los avances obtenidos al presente en a obtenciOn de tejido embriogénico y el desarrollo do plantas a partir de este tejido, permiten disponer do un protocolo do propagaciôn a nivel experimental do embriogénesis somática que necesita ser adecuado a los genotipos de intorés asi como perfeccionado en sus diferentes etapas a fin de quo pueda ser aplicado a Ia escala camercial. Tomando en cuonta los puntas antoriores, en esta comunicaciOn se presentan resultados obtonidos en Ia extrapolacion a genotipos de interés comercial; un rnejoramiento do Ia inducciOn do tejido embriogénico primario, asi coma el desarrollo de embriones y conversion a plántulas.
Materiales y métodos
Material vegetal
Protocolo experimental
La esterilizacian tie los medios de cultivo fue realizada en autoclaves de presión do vapor a 12°C durante 15 mm Una vez sembrados los explantos, los cultivos fueron incubados en salas do cultivo en condiciones do obscuridad y a una temperatura media de 25°C.
Factores estudiados
Las variables do respuesta cuantificadas on osta investigaciOn fueron:
Las lecturas para toma de datos fueron realizadas entre Ia sexta y octava semana después de Ia puosta en cultivo. Todas las observaciones y tomas do datos so realizaron con Ia ayuda de un microscopio estereoscopio a 2,5 X.
Los tratarnientos fuoron distribuidos en base a un diseno completamente al azar, donde cada unidad experimental estuvo ropresentada por una caja do Petri do 5,5 cm de diametro con dos botones florales; cada uno de los tratamientos estuvo representado por 10 repoticiones. La oficioncia de cada tratamiento fue evaluada mediante Ia prueba do independoncia do medias de Chi cuadrado al 5% (Dagnelie 1975).
Resultados y discusiôn
Importancia de Ia fuente de carbono
En el genotipo H 20, Ia mejor induccion do tejido con aptitud embriogénica se obtuvo on los tratamientos de sacarosa 60g/l (12,8%), sacarosa 80g11 (16,7%), glucosa 60g11 (12,7%) y glucosa 50g/I con un porcentaje do 16,9% do induccion de tejido embriogAnico. En el genotipo RIM 88, Ia mejor induccion fué cuantificada cuando el medio se complementó con glucosa a 50 y 60g/l y sacarosa a 80g/I con respuostas tie 2,1%, 1,0% y 2,0%. La respuesta més baja, fue observada en los tratamientos a base de maltosa on sus tros concentraciones.
Por otra parte, so observô que en ambos genotipos, tanto Ia sacarosa como Ia glucosa a las concentraciones ovaluadas; provocaron una fuerte callogenesis primaria; miontras quo Ia maltosa en concentraciones do 40 y 80g/l no indujo ninguna callogénesis.
En lo que corresponde al porcentaje tie explantes muertos se cuantifico quo Ia mayor cantidad de mortalidad ocurriO con el tratamiento maltosa en sus tres concentraciones, obteniendose por ol contrario el menor porcentaje do mortalidad do explantes con glucosa a SOy 60g/l ambos con un porcentaje tie 3,0%.
Efecto do Ia composicion nutricional del medio do cultivo
En cuanto a Ia induccián de callos ombriogénicos, los resultados obtenidos presontados en a Cuadro 3 muostran quo en todos los modios evaluados fué posible Ia obtenciôn do callos embriogénicos, independientemente del genotipo del explanto. En el caso del genotipo H 20 Ia respuesta cuantifucada resultó similar en los medios MS-2, MS-3 y MS-4; on el caso del don RIM 56 Ia mejor induccion do ombriogénesis Se observô a partir de explantes cultivados en los medios MS-3 y MS-4. En ambos genotipos so observa que ol medio MS-iS quo corresponde a a formulacion original do Murashige y Skoog, rosulta poco favorable para Ia inducción do embriogenesis.
Importancia do Ia composición hormonal del medio de inducción
En cuanto a Ia reacciôn de los explantes del genotipo H 20, las mejoros tasas de induccion fueron obtonidas con Ia combinaciôn 2,4-0 con kinetina y 2iP, y 2,4,5-T con 2iP: en el caso del don H-31 Ia combinaciôn del 2,4-0 con las citoquuninas resultó en un efecto similar. En cuanto a Ia auxina 2,4,5-T a mejor rospuesta fue observada cuando esta se combinô con 2iP.
Cuando Ia auxina 2,4-D fue utilizada, Ia mejor respuesta tue cuantifucada cuando esta so complementO con Ia kinetina; en el caso especifico del don H-20 se observô un 58% do inducciOn de tejido embriogenico. De acuerdo a Ia respuesta obtenida en el tratamiento testigo, se observa que los explantes no mostraron en este tratamiento ningun desarrollo ni reaccion de embriogenesis. Los resultados anteriores indican entonces quo so requiere do Ia presencia de los reguladores de crecimionto en el medio de cultivo para a obtonción do tojido con aptitud embriogenica.
Sobre Ia base do estos rosultados y con Ia finalidad do detorminar el efecto hormonal en Ia fase de expresion, es decir en el medio secundario de embriogenesis, so evaluo el efecto do las citoquininas kinetina y BAP en Ia oxpresión do Ia embriogénesis; como testigo so utilizO el medio nutritivo sin ningun regulador de crecimiento.
En el Cuadro 5 son presentados los resultados obtenidos de esta prueba, en donde so observa quo indopondiontomente del medio primario, Ia prosencia de citoquininas en el medio secundario no es nedesaria para Ia obtencion de tejido embriogénico; y segUn el genotipo del tejido cuitivado las citoquininas indujeron efectos negativos en Ia expresiôn do Ia embriogenesis.
Las proporciones cuantificadas en Ia produccion do callos ombriogenicos en los diversos modios resultaron variablos y no muestran una tendoncia generalizada hacia el mejoramiento de Ia respuosta en ombriogénesis y en el caso particular del don H 31 Ia rospuesta cuantificada en dasi todas las situaciones resultó inferior a Ia cuantifucada en el testigo sin ninguna citoqtjinina.
Efecto do pretratamientos de botones florales a bajas temperaturas
La incubacion de botones florales a 5°C resulto en una mejora de la induccion embriogenica cunando estos fueron tratados durante 30 y 120 min, el resto de los tratamientos dio como resultado valores in feriores al testigo
Respuesta de 12 genotipos a Ia induccion do embriogenesis
Los resultados obtenidos se prosentan en el Cuadro 7 donde se obsorva que Ia induccon de callos ombriogenicos tue posible en todos los gonotipos estudiados. Destacan por Ia mayor proporciOn do callos embriogénicos los genotipos RIM 68 y RIM 56; el RIM 88 presonta una rospuesta considorada mntermedia y Ia respuesta mas baja tue observada on los clones H 31 y H 16. Es de hacor notar quo los clones RIM 68 y RIM 76A pertenecen al tipo gonético Criollo y quo se consideran dificiles a la propagacion horticola convencional por enraizado do estacas o injertado.
Las tasas de explantes ombriogenicos obtenidos en este estudio son comparables a los reportados por Lopez et al. (1993) y Lopez (1994), quien reportO valores comprendidos entre 1,3 y 18,0%, y a los obtenidos por Alomanno (1995) quien reporta una mnducciOn del 4,9 al 11,6% do embriogénesis. Es necesario precisar que los estudios anteriormonte citados se desarrollaron utilizando explantes do genotipos Forasteros.
Dentro del grupo de genotipos evaluados por Alemanno (1995) se incluyeron los clones Mexicanos RIM 43 y RIM 106 en los cuales no tue posible Ia inducciOn do tejido embriogénico, por lo quo fueron clasificados por esto autor como recalcitrantes a a embriogenesis somática
En este estudio, el protocolo desarrollado fue aplicado al cultivo de explantes de 12 clones Mexicanos (RIM 24, RIM 88, RIM 56, RIM 68, RIM 76A) los cuales reaccionaron positivamente obtoniéndose callos embriogénicos en todos los genotipos, haciAndoso notar que los tres primeros pertenecen al grupo genético Trinitario y los dos Cjltimos al grupo genético Criollo.
El éxito de a embriogénesis somática in vitro como via de propagacion esta grandemente influenciado, entre otros factores por a naturaleza del medio do cultivo empleado, del efecto do los reguladores del crecimionto y dol genotipo.
Asi, durante las fases do induccion y expresiOn de Ia ombriogénosis diferentes secuencias de medios do cultivos puoden sor necesarmos, acorde con los requorimientos nutricionales de cada especie y de los tejidos o células en cultivo (Ammirato 1989 y 1993).
EI medio do cultivo contiene basicamente: las sales minerales ciasifmcadas como macro y microelementos, las substancias orgánicas (vitaminas, azücares y aminoacidos), los reguladores do crecimiento y los productos complejos como Ia caseina hidrolizada, el extracto de malta, el agua do coco, y el carbon activado. Eventualmente otros productos puodon ser agregados al media, como por ejemplo antioxidantes 0 inhibmdores do accion do efectos toxicos generados por moleculas 0 compuestos que son excretadas por las mismas celulas,
El media de Murashige y Skoog MS’ ha sido ampliamente reportaclo para Ia embriogénesis on numerosas especies vegetales; y fué reportado inicialmente por Lopez et a/. (1993) y Alemanno (1995) como adecuado para el proceso de embriogenesis en cacao, los resultados obtenidos en este trabajo confirman estos reportes. Sin embargo, dada Ia diversidad de genotipos seleccionados en diversos centros do investigaciOn del mundo, os innegable que para Ia mnduccian do embriogenosis somática on otros genotipos, se requeriran do adaptaciones al mismo, especialmente para el cultivo do explantes do genotmpos considerados recalcitrantes.
En base a ía totipotencialidad do a céfula vegetal, todas as partes de una planta son aptas a formar ombriones somatmcos, sin embargo, en Ia prăctica se ha demostrado quo los tejidos más aclecuados son en general los niăs juveniles y menos diferenciados (Ammirato 1993; Carman 1990); aunque también ha sido reportado el efecto inductor do protratamientos a los tojidos antes do la siembra.
La obtenciOn do tejidos embriogénicos a partir de piezas florales ha sido reportada en varias especies como por ejemplo: 1-lemerocallis citrina (Zhou et a!. 1984), Secale cereale (Linacoro y Vazquez 1990), Punica granaturn (Mahishi 1992).
En el caso espocifico del cacao, el potencial embrmogenico de las piezas florales (petalos, estaminodios y filamentos de anteras) como explante inicial para Ia formacion de Ia embriogénesis, ya habia sido reportado anteriormente por (Lopez et a!. 1993; LOpez 1994 y 1996) y Alomanno (1995) el cual es confirmado por los valores registrados en las tasas do induccion do tejido embriogenico obtenidos en este estudio.
Por otra parte, los reguladores del crecimionto desempenan un papel determinante en Ia inducciOn de Ia embriogénesis, destacando el tipo de horniona, Ia concentracmon en el media de cultivo y Ia duraciOn de Ia fase de inducciOn. Estos reguladores actüan do manera especifica sobre el comportamiento celular, como por ojemplo, Ia estimulaciOn do Ia divisiOn celular y Ia orientaciOn do las celulas hacia Ia dosdiferenciaciOn. Do manera general eI proceso de embriogénosis se inicma en un modio do cultivo quo contione principalmente auxinas y citocininas, solas o conibinadas.
Lopez et at (1993), Lopez (1994) y Alomanno (1995) reportan que a embriogenosis somática a partir de piezas florales do cacao os posible on un medio enriquocido con el balance hormonal 2,4-0 (1mg/I) + kinetina (0,25mg/I), datos quo so confirman con los cuaritificados en esta investigacion. Aunque es de hacer notar quo en esta investigaciOn so reporta par primera vez el efecto positivo sabre Ia inducción do embrmogenesis de Ia auxina 2,4,5-T en concentracion do 1mg/I, no existén reportes previos quo indiquen Ia aplicacion do este regulador del crecimiento on Ia inducciOn do embriogénosis somática en Theobroma cacao.
En cuanto al ofecto inductor do bajas temperaturas en Ia embriogenesis a partir de tejidos soniáticos, este ha sido reportado on diversas especies, es importante hacer notar quo no existen reportes bibliograficos previos en cacao quo permitan establecer una comparaciôn con los rosultados obtenidos en osth invostigaciOn No obstante, los resultados observados ponen en evidencia el efecto mejorador do Ia embriogénosis do pretratamientos de los botones florales antes de Ia puesta en cultivo a temporaturas do 0 y 5 grados contigrados
Debido a Ia importancia do los reguiadores do crecimiento en el proceso de ombriogenesis, se confirma a necesidad do estos en el medio de cultivo inicmal para inducir Ia reacciOn embriogénica; sin embargo en el medio secundario do expresiOn las citoqumninas no son necesarias. Lo anterior indica que para quo eI procoso de embriogenesis se desarrolle es necesario un protocolo do inducciOn — expresiOn, en el cual los reguladores de crecimiento solo estarian presentes en el medio primario para provocar Ia inducciOn, posteriormente las hormonas so suprimen para favorecer Ia oxpresiôn. La eficioncia de oste protocolo es corroborada por a buona calidad de Ia embriogenesis obtenida como so aprecia en Ia Figura 1 ya quo los embriones obtenidos fueron capacos do desarrollarse hasta plantas en condiciones de invemadero, lo cual indica entonces que estos tienen a capacidad para el desarrollo posterior sin que ocurra ninguna malformaciOn ni el bloqueo de estos. El crecimiento observado en las plantas regeneradas por esta via es similar al do una planta originada do semilla.
Protocolos similares de inducciOn — expresiOn han sido reportados en diversas especies entre las que es posible citar Ia alfalfa Medicago sativa (Walker y Sato 1981), Ia zanahoria Daucus carota (Gorst et a!. I 987) y Ia soya Glycine max (Parrot et a!. 1988), en las cuales unas pocas semanas de cultivo en un medio complementado con hormonas es suficiente para inducir Ia reacciOn de embriogénesis. Un cultivo prolongado de los explantes en presencia de hormonas provoca una monogenesis anormal que se traduce en malformaciones y bloqueos en el desarrollo posterior de los embriones obtenidos.
De acuordo con Lopez et at (1997a) los clones de Ia sorie RIM soleccionados en eI CE Rosario Izapa, constituyen una población bastante homogenea y omparentada, por lo que puede preveerse una respuesta similar en estos materialos.
Este estudio muostra entŕncos, que adaptaciones a Ia composicion nutricional y hormonal del modlo desarrollado por Lopez et a!. (1993) rosultaron favorables para Ia obtenciôn de tojido embriogenico en genotipos considerados roclacitrantes. Sm bien os cierto que Ia respuesta embriogénica os variable segUn el genotipo, los resultados obtenidos indican Ia factibilidad do aplicación de osta via do propagacian a clones do intorés industrial por su alta calidad como son los gonotipos Crmollos, que en general son considerados de dmficil propagacion por las tecnicas horticolas convencionales (Enriquoz 1985; Loon 1987; Flores y Vera 1995).
Sin embargo, oste estudio no pormito concluir de manora definitiva a corca do Ia respuosta embriogenica y el gonotipo del material (Criollos, Trinitarios y Forasteros) definidos on cacao (Cope 1984; Leon 1987), los resultados obtenidos son indicadores de que el potencial embriogenico existe on genotipos de ostos grupos genéticos y quo para desarrollar este se requieron adaptaciones 0 adecuaciones al protocolo.
Potencial de Ia propagación de Theobroma cacao por embriogenesis somatica
La propagaciOn do plantas do cacao es una limitante a nivol mundial por lo que las investmgacionos quo so desarrollan en diferentes partes del mundo so encammnan hacia el perfoccionamiento do las técnicas do clonamionto ya disponiblos como son: ol injortado convencional, el enraizado de ramas, ol microinjortado en plântula y las nuovas tecnologlas de cultivo do tojidos como Ia embriogenosis somatica (Vow y Lim 1994; Flores y Vera 1995; Lopez 1996; Furtek et all 1994).
Debido a que Las técnicas disponibles de propagaciOn clonal no son suflcientes para cubrir Ia domanda do plantas requoridas para poder renovar las plantaciones, como ejomplo se tione quo para el caso del ostado de Chiapas en Mexico so necesitan 40 000 000 do plantas para ronovar las 30 000 ha que oxisten, a una densidad de plantado de 1100 árboles por ha, de manera que los estudios hacia Ia optimizaciOn del procoso de embriogAnesms somatica han cobrado mayor interés.
Los rosultados obtonidos en oste trabajo de investmgacian indican, quo Ia inducciOn de Ia embriogenesis somatica es factible dosarrollarla on genotipos denominados recalcmtrantes, y quo ademas tiene aplicacion inmediata para poder optimizar el procoso.
Tomando en cuenta ol potoncmal quo ofreco esta tecnica hay nocesidad do perfoccionarla para poder aplicarla a Ia oscala comercial.
La ombriogenesis somâtica ya desarrollada permitiria aplicar otras técnicas do biotocnologia como son Ia ingonioria gonética y Ia fusiOn do protoplastos, tecnicas que permitirian desarrollar en un tiempo rolativamonte corto (3 —4 anos) nuevos clones, a diforencia del mojoramiento convencional quo requiere do 20 a 30 ańos para obtonerlas,
La optimizaciOn dol proceso do ombriogénesis somatica en medio liquido, por ejemplo on bioroactoros, serâ do gran importancia econOmica ya quo permitira Ia producciOn a gran escala y con un costo roducido do grandos cantidades do plantas de una variodad dotorminada. Estas plantas podrán ser utilizadas de manora directa para ol establocmmiento de cultivos 0 bion conservarso.
Conclusiones y perspectivas
La composmcion mineral del medio MS-4, quo corrosponde a una modificafiOn de Ia formulacion original, resultO Ia mas adecuada para dosarrollar el proceso do ombriogénosis somática a partir do oxplantos do piezas floralos de cacao en gonotipos seloccionados en Mexico.
Las composicionos hormonales a base de 2,4,5-1 (1mg/I) o 2,4-D (1mg/I) y kinetina (0,25mg/I) en el medio primario, rosultaron las mas adocuadas para Ia induccián do tejido embriogénico. La glucosa en concontracionos do 50 y 60g/l y sacarosa a 80g/l resultaron las fuentos de carbono mas adocuadas para Ia inducciOn de explantes embriogénicos. En el medio secundario de expresian do Ia embriogenesis no es nocesaria Ia presencia do citocininas para Ia obtencion de tejido ombriogenico. El pretratamionto de botonos florales antes do Ia siembra a temperatura do 0°C durante 15 y 20 mm, 0 a 5°C duranto 120 mm inducen una mejora on Ia proporcián do tejido ombriogénico formado.
Los rosultados obtonidos permitieron intograr modificaciones al protocolo do ombriogénesis previamonte desarrollado, el cual mostrO su oficiencia ya quo fue posible a mnduccion do tojido embriogénico on explantes de 12 genotipos, algunos de ellos considerados rocalcitrantos.
Los resultados obtenidos indicarian quo Ia obtencion do tejido con aptitud ombriogénica es una resultante de a mnteracciOn del genotipo del explante, dol balance hormonal y de a composician mineral y energetica presonte en el medio primarlo do mnducción.
Bibliografla
Adu-Ampornah V., F.J. Novak, R. Afza, M. Van Duren y M. Perea-Dallos. 1988. Initiation and growth of somatic embryos of cocoa (Theobmma cacao L.). Café Cacao The 32: 187—1 99.
![]()

Biotech Glossary |
Bioinformatics |
Lab Protocol |
Notes |
Malaysia University |
Avanzos en Propagación de Cacao - Theobroma cacao - por Embriogenesis Somática en Mexico.
Orlando López-Baez, Jose Luis Moreno-Marginez y Sonia Pacheco-Rodas
Facultad De Ciencias Agricolas, Universidad Autonoma De Chiapas, Apartado Postal 34, Tapachula, Chiapas, 30700 Mexico
Corno material vegetal para el estudio Se utilizaron botones florales do dos clonos por cada experimonto, posteriormente tomando en cuenta los resultados obtenidos en los diversos ensayos, so hizo una evaluación do genotipos considerando los clones siguientes: RIM-24, RIM-44, RIM-56, RIM-88, RIM-105, H-12, H-13, H-16, H-20, H-31, seleccionados para ol estado do Chiapas.
La inducciOn do tejido embriogénico Se realizô a partir do explantos do piezas florales (pétalos, estaminodios y filamentos de anteras) tomados de botones antes de Ia apertura do acuerdo al protocolo técnico desarrollado por LOpez (1994). Los botones florales fueron desinfectados por inmorsiôn on cloralex al 80% durante 13 mm y enjuagados tres veces en agua destilada estéril. Una vez ostorilizados, los botones fiorales fueron disectados y los explantes (pétalos, filamentos de las anteras y ostaminodios) puestos en cultivo en cajas de Petri de 5,5 cm do diâmetro que contenian los medios do cultivo con los diferentes tratamientos de estudio. Como medio bâsico nutricional so utilizO Ia formulaciOn del medio denominado “MS” y “MS’ moditicado quo corresponde a una modificación en Ia concentraciOn tie los macroelomontos y microelemontos del medio do Murashige y Skoog (Ammirato 1989; Carman 1990), complemontado con glucosa Sag/I, golrite 2,5g/I y ol pH ajustado a 5,5 Anexo 1).
Los factores de estudio evaluados fueron:
En ol Cuadro 1 so presentan los resultados obtenidos do Ia ovaluaciOn de tres fuentes do carbono a tres concentraciones, tomando como referoncia Ia reaccion do oxplantes do los genotipos H 20 y RIM 86.
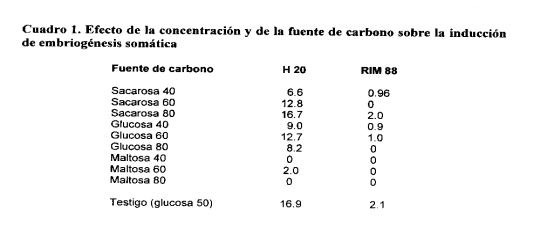
En el Cuadro 2 se presenta Ia respuesta cuantificada en induccián de callogenesis primaria y a mortabdad de explantes de los genotipos H 20 y RIM 56 segUn cuatro medios de cultivo (compasiciOn indicada en el Anexo 1). Se observa quo Ia callogenesis es generalizada y similar en los modios evaluados, Una respuesta similar en ambos genotipos se observa en a variable mortalidad de explantes al inicio del cultivo. De manera general Se observa quo a menor tasa de mortalidad do explantes ocurre en ol medio MS-4.
.jpg)
.jpg)
En el Cuadro 4 so presentan los resultados obtonidos do Ia evaluacián do las auxinas 2,4-D y 2,4,5-T evaluadas en combinaciôn con las citoquininas kinetina y 2iP on explantes do los genotipos H-20 y H-31. Se observa que todas las combinaciones hormonalos provocaron un ofecto de induccion de callos embriogénicos. Los oxplantos cultivados en un medio sin reguladores do crecimiento no formaron en ningün caso callos embriogénicos.

Con Ia finalidad do determinar el efecto do pretratamientos de los botones florales a bajas temperaturas, previo a Ia siembra, so evaluaron diferentos periodos do incubación a las temporaturas do 0 y 5°C. SegUn puede observarso en los rosultados presentados on el Cuadro 6, Ia incubacion a 0°C muostra un efecto positivo sobre Ia inducción de Ia ombriogenesis cuando los botones son pretratados por un tiempo do IS y 20 mm, ya que so obsorva quo Ia proporción do callos embriogenicos es suporior a Ia cuantifucada on el testigo. Tratamientos do mayor duración en tiempo no rosultaron favorables a Ia embriogénesis.
.jpg)
.jpg)
Tomando como base los resultados obtenidos en los ensayos presentados anteriorniente so realizaron modificacionos al protocolo de induccion, y postoriormente fue aplicado al cultivo de oxplantes do 12 clones algunos de ellos considorados rocalcitrantos a Ia propagación in vitro (Lopez 1996; Lopez et al 1997a).

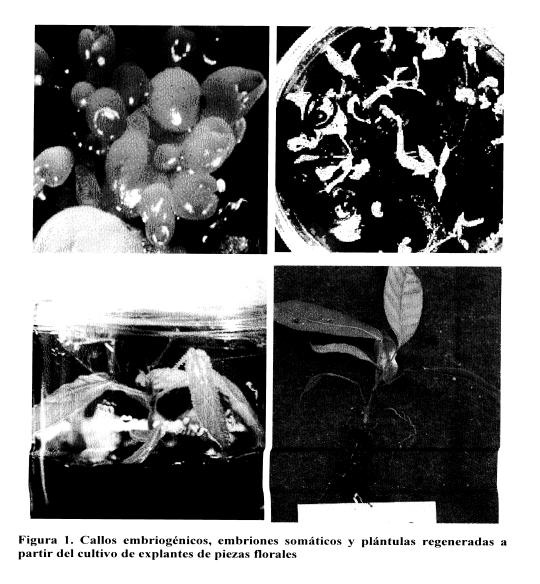
La forma do perpetuarse do el cultivo do cacao Cs mediante Ia propagacion ya sea sexual o asexual (Enriquez 1985; Loon 1987). Sin embargo, dobido a Ia naturaleza alogama del cultivo no so rocomionda realizarse via sexual ya quo las plantas obtenidas presentan alta variacion genetica.
Aguilar ME., M.V. Villalobos y N. Vazquez. 1992. Production of cocoa plants (Theobroma cacao L.) via micrografting of somatic embryos. In vitro Cell. Dev. Biol. 28: 15-41.
Alemanno L. 1995. Embryogenese somatique du cacaoyer Theobroma cacao L. Contramntes et perspectives. These de Docteur en Sciences, Université de Montpellier II, Sciences et Techniques du Languedoc. 193 pp.
Ammirato P.V. 1989. Recent progress in somatic embriogenesis. Int. Assoc. for Plant Tissue Culture Newsletter 57: 2-16.
Ammmrato P.V. 1993. Embryogenesms. Pagmnas 82-123 in Handbook of plant cell culture. Techniques for propagation and breeding. Vol I. (Evans, D. A, Sharp, W. R., Ammmrato, P. and Yamada, V., eds). McMiIlan, New York, United States of America.
Carman J.G. 1990. Embryogenic cells in plant tissue cultures: ocurrence and behaviour. In Vitro Cell. Dev. Biol. 26: 746-753.
Chatelet P. N. Mmchaux-Ferriere y P. Dublin. 1992. Potentialites embryogenes du nucelle du tégument interne de graines immatures de cacaoyer (Theobroma cacao L.) Compte-Rendus de l’Académie des Scmences. Parms 315: 55-62.
Cope F. W. 1984. Cacao Theobrnma cacao L. (Sterculiaceae). In Simmons N. W. (ed). Evolutmon of crop plants. Longman, London.
Dagnelie P. 1975. Théorie et methodes statistmques: apprications agronommques. Les Presses Agronomiques de Gembloux, Gembloux, Belgmque. 463 pp.
Enriquez GA. 1985. Curso sobre el cultivo del cacao. Turrialba, Costa Rica, CATIE. Serie materiales de enseńanza n°. 22. 239p.
Esan E.B. 1977. Tmssue culture studies on cocoa (Theobmina cacao L.) A supplementation of current research. Paginas 116-125 in Actes de Ia 5a. Conference Internationale sur Ia Recherche Cacaoyére, September 1-9, 1975, Ibadan, Nigeria.
Flores F. y B.J. Vera. 1995. Influencia de Ia fenologia sobre el enraisamiento de ramillas y prendimiento de mnjertos en clones de cacao. El Cacaotero Colombiano 14: 26—34.
Furtek 0., Anderbran T., Chunyan G., Glazier B., Pate1 V., Samn S., Simka H. y Traore A. 1994. Applmcations of biotechnology for cocoa tree improvement. Paginas 11-12 in Malaysian Internatmonal Cocoa Conference. 20-21 october, Kuala Lumpur, Malaysia. Malaysia Cocoa Board. Extended Abstracts.
Gorst J., Overall R. L. y Wemeke. 1987. Ionic currents traversing cell cluster from carrot suspension cultures reveal perpetuation of morphgenetmc potential as distinct from induction of embryogenesms. Cell Differentiation 21: 101-109.
Kononowicz H., AK Kononowicz y J. Janick. 1984. Asexual embryogenesis via callus of Theobroma cacao L. Zemtschrift für Pflanzenphysiologie 113: 347-358.
Leon J. 1987. Pagmnas 337 —343 in Botanica de los cultivos tropicales. IICA, San José, Costa Rica.
Linacero R. y A.M. Vazquez. 1990. Somatic embryogenesms from mnmature infirescences of rye. Plant Science 72: 253—258.
LiztR.E. 1986. Tissue culture studies with Theobroma cacao. Paginas 111-120 in Proceedings of the Cacao Biotechnology Symposium. (Dmmick, P.S., ed). Pennsylvania State University Press, University Park, Pa, United States of America.
Litz RE. 1987. Application of tissue culture to tropical fruits. Paginas 407-418 in: Plant tissue and cell culture. (Green, C. E. eta!., eds). Alan R. Liss, New York, United Kingdom.
Lopez B.C. 1994. Embryogenese somatmque et regeneration de plantes de cacaoyer Theobroma cacao L. a partir de pieces florales. These de Docteur en Scmences, Ecole Nationale Supérieure Agronomique de Rennes, France. 137 pp.
Lopez B.O. 1996. Biotecnologias aplicadas al mejoramiento genético y propagacian del cacao. Paginas 176-179 in: Actas de Ia ix ReuniOn Cientifica-TecnolOgica Forestal y Agropecuaria, 5-6 de Dmcmembre, Villahermosa, Tabasco, Mexico.
LOpez B.O., H. Bollon, A.B. Eskes y V. Petiard V. 1993. Embryogenése somatique de cacaoyer Theobroma cacao L., a partir de pieces florales. Compte-Rendus de lAcademie des Sciences 316: 579-584,
Lopez B.C., H. BolIan, M. Alvarez y V. Petiard. 1996. Ex-vmtro performance and nuclear DNA ploidy of cocoa plants regenerated by somatic embryogenesms. Paginas 589-594 in Proceedings of the 12th, International Cocoa Research Conference, November 17-23, 1996, Salvador, Bahia, Brazil.
Lopez B.O., G.M. Esponda, yB. Hernandez, V.G. Fraire, H.M. Evans y A. Fontanel. 1997a. Les biotechnologies appliquees a ‘amelioration genetique et Ia propagation du cacaoyer au Mexique. Paginas 8-9 in Résumés Viemes Journees Scientifiques du Réseau Biotecnoiogies-Genme Genétique des plantes de I’APELF.UREF. Universmte de Paris Sud,Centre Universitaire d’Orsay, Francia. 30 junio al 3 de julio de 1997.
Lopez B.O., G.M. Esponda, V.B. Hernandez, V.G. Fraire, H.M. Evans y A. Fontanel. 1997b. Progresos recmentes en el clonamiento del cacao. Paginas 15-16 in Actas del Primer Congreso Venezolano del Cacao y su Industria, 18 al 21 de noviembre 1997, Maracay, Aragua, Venezuela.
Mahishi D.M. 1992. Regeneration of petal callus in Punica granatum. Pagina 39 in Advances in gene technology: feeding the world in the 21M century. Short reports Vol 1. (%Mielan, W. J. eta!., ed). Miami, Florida, United States of America.
Parrot WA.., G. Dryden, S. Vogt, OF. Hmldebrand, GB. Collins y E.G. Williams. 1988. Optimization of somatmc embryogenesis and embryo germination in soybean. In Vitro Cell. Dev. Biol. 24: 817—820.
Pence V.C. 1989. Cacao (Theobrorna cacao L.). Paginas 203-221 in: Biothecnology in Agrmculture and Forestry. Vol 5. Trees II. (Bajaj, Y.P.S., ed). Springer-Verlag, Berlin,
Petiard V., J.P, Ducos, B. Florin, C. Lecouteux, H. Tessereau y A. Zamarripa. 1993. Production en masse et conservation des embryons somatiques. Pagmnas 175-191 in Vol. I Proceedings of the 4eme Rencontres Internationales sur es Semences. Aspects Fondamentaux et Appliqués de Ia Biologie de Semences, Angers, France, 1992. (Come D. et Corbineau F., eds).
Sondhal M. R., T.H.B. Sereduk, CM. Bellato y Z. Chen. 1998. Somatic embryogenesms and plant regeneration of cacao. European Patent Application, Publication 0293598 A2. Munchen, Germany. llpp.
Yow ST. y DIlL. Lim. 1994. Green-patch budding on very young cocoa rootstocks and side grafting of mature trees. Cocoa Growe(s Bulletin 47: 27—41.
Walker K. A. y S.J. Sato. 1981. Morphogenesis mn callus tissue of Medicago sativa: the role of ammonium ion on somatic embryogenesis. Plant Cell, Tissue and Organ Culture 1: 109—121.
Zhou P., J. I-lu y H. Fan. 1984. Studies on inducing embryomd and plant from stamens and unripe seeds of long yellow day-lily in vitro. Pagina 72 in Abstracts International Symposium on Genetic Manipulation in Crops. Beijing, China, 1984.