Biotechnology can provide a powerful set of tools for improving the yield of cocoa trees and the quality of beans. Basic tools encompass tissue culture, genetics, and molecular biology. Some of the benefits for cocoa improvement that can be provided by biotechnology include.
Propagation by somatic embryo genesis
Objective
Materials and methods
Secondary somatic embryogenesis. Primary somatic embryos were placed on expression media containing a cytokinin for several weeks, then transferred to similar media without growth regulators.
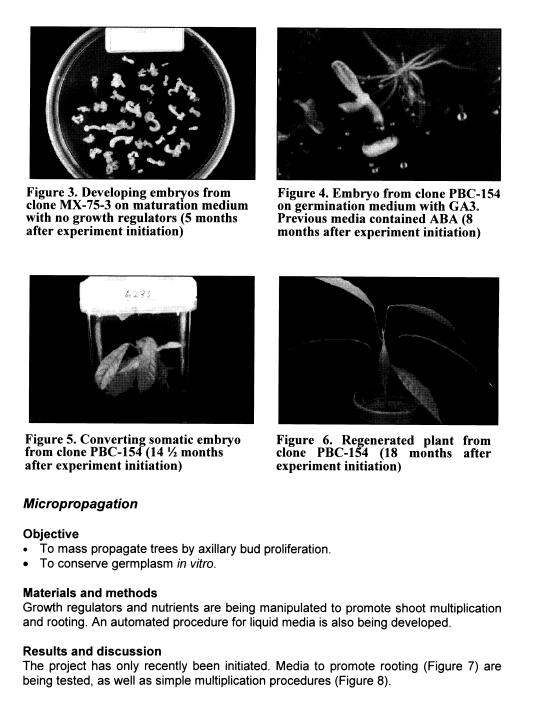
Embryo maturation. Embryos were allowed to grow on maturation media containing minerals, sucrose, and vitamins until they were developed enough to transfer to conversion media. Maturing embryos have also been transferred to similar media containing ABA for 3 - 6 weeks.
Embryo germination. Large embryos were placed on germination media containing GA3 for 3 - 6 weeks.
Embryo conversion. After embryos produced primary and sometimes secondary roots and are several centimeters long, they were transferred to conversion media with reduced salt concentrations, sucrose, and vitamins and placed in the light room.
Results and discussion
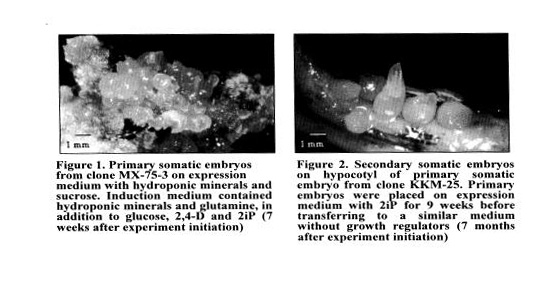
Secondary somatic embryogenesis. After several weeks on minimal media (sugar, minerals, vitamins) containing 0.1 mg/I 2iP and several more weeks on minimal media with no growth regulators, secondary somatic embryos appear (Figure 2).
Embryo maturation and germination. ABA in maturation media along with GA3 in germination media improves embryo rooting (Figures 3 and 4).
Embryo conversion. After the maturation and germination phases, when embryos have produced at least primary roots and are several centimetres long, they are transferred to media containing a reduced mineral concentration, sucrose, and vitamins and placed in the light room in order to stimulate development of true leaves (conversion into plantlets). Problems were encountered, however. Hypocotyls often turned brown and embryos failed to develop leaves. Embryos persisted for several months without showing any sign of further development. Most eventually died.
Several changes were subsequently made in conversion media and culture conditions. The gelling agent was changed from Phytagel to agarose because Phytagel binds bivalent cations such as calcium. Lights were changed from sunlight-balanced to a combination of cool-white fluorescent and incandescent. Light intensity was reduced and the temperature in the light room was lowered by a few degrees, to 24 — 25° C.
We have not investigated any of these changes individually. However, our conversion frequencies have improved considerably. Thirty-sIx (36) somatic embryos began to convert into plantlets since these changes were made. Seventeen (17) were discarded, usually due to contamination. Of the 19 remaining ones, 5 are producing normal true leaves (Figure 5), 5 have been transferred to pots (Figure 6)
Abscission zone formation in pods
Objectives
Results
Acknowledgements
The authors would like to thank the Director General of the Malaysian Cocoa Board for his permission to publish this paper The excellent technical assistance by Mr. Heden Jainuddin, Mrs. Mavis Peter Jaus, Mrs. Sairan Asim, Mrs. Siti Salmiah Mohd. Sadan, Miss Rafiah Hf Karim, Mr Willy Kimsui, Mr. Henipa Bin Jambol, and Mr. Masri is greatly appreciated. These projects are mainly funded by the Intensive Research Priority Area (IRPA) programme of the Ministry of Science, Technology and Environment (MOSTE), Malaysia.
References
Alemanno L., M Berthouly and N. Michaux-Ferriere. 1997. A comparison between Theobroma cacao L. zygotic embryogenesis and somatic embryogenesis from floral explants. In Vitro Cell. & Develop. Biology. Plant 33: 163-172![]()

Biotech Glossary |
Bioinformatics |
Lab Protocol |
Notes |
Malaysia University |
Biotechnology Research by the Malaysian Cocoa Board
Douglas Funek , Rosmin Kasran, Tan Chia Lock, Lea Johnsiul, AzHar Mohammed, Victor Hartney, Meriam Mohd. Yusof, Kelvin LaMin and Lee Ming Tong
Malaysian Cocoa Board. Wisma Sedco. Locked Bag 211 88999 Kota Kinabalu. Sabah.
Malaysia (http://www.koko.gov.my: *dbf@pc.jarjng.mv)
Primary somatic embryo genesis. Staminodes — often with the base of filaments attached - from large but unopened flower buds were placed on induction media containing either DKW minerals or one derived from a cocoa hydroponic mix, glucose, MS vitamins, organic supplements, 1.0-2.0 mgIl 24-0, and 0.1 mg/I 2iP. After 3 - 4 weeks, calli were transferred to expression media containing minerals, sucrose or maltose, MS vitamins, and organic supplements, but no growth regulators.
Primary somatic embryogenesis. Overall the two most important factors influencing embryogenesis frequencies are the cocoa ckne and the induction media. Regardless of the medium, staminodes from some clones either produce abundant callus or grow poorly. In either case, few or no embryos are produced. The only organic supplement we have found that may promote embryogenesis is glutamine in the induction media (Figure 1). Other supplements such as coconut water, malt extract, and yeast extract often either lowered embryogenesis frequencies or resulted in translucent and deformed embryos. Sucrose and maltose in the expression media resulted in approximately the same embryogenesis frequencies
Results
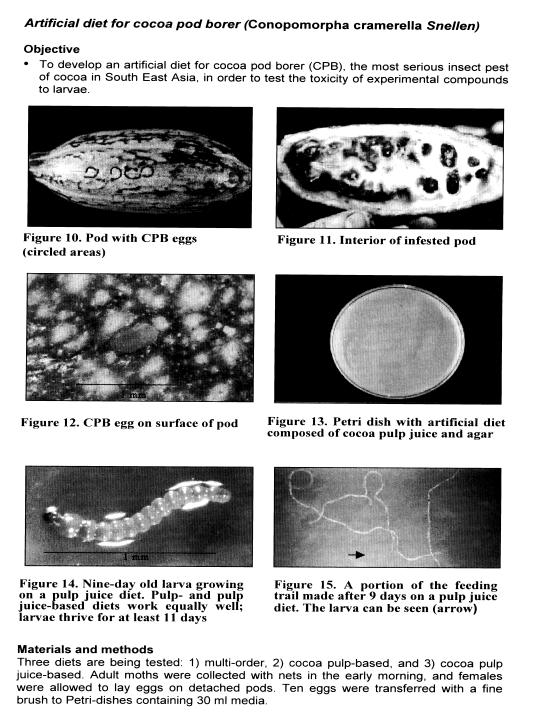
A diet composed of cocoa pulp juice or cocoa pulp sustains CPB larvae at least 11 days (Figures 10 to 1 5). Experiments were stopped afterwards due to contamination.
This is a new project. RNA is still being extracted from cocoa pods, flowers, and other tissues. PCR primers have been synthesised for the cellulase and jointless genes. Although the jointless PCR primers were designed from the tomato gene. a single band has been produced with cocoa DNA using moderately stringent conditions. This band will be sequenced, and if it is the cocoa jointless homologue it will be used as a probe to isolate the corresponding cocoa gena
Li Z., A. Traore, S. Maximova and M. I Guiltinan. 1998 Somatic embryogenesis and plant regeneration from floral explants of cacao (Theobroma cacao) using thidiazuron. In Vitro Cell. & Develop. Biology. Plant 34: 293-299
Lopez-Baez 0., H. Bolton, A. Eskes and V. Petiard 1993. Somatic embryogenesis and plant regeneration from flower parts of cocoa Theobroma cacao L. Comp Rendu Acad Sci. Ill -Life Sciences 316: 579-584
Mao L., D. Begum. H.W Chuang. MA. Budiman, E.J Szymkowiak, E.E. lrLsh and R.A. Wing. 2000. Jointless is a MADS-box gene controlling tomato flower abscission zone development. Nature 406: 910-913.
Singh, P. and RE. Moore (eds). 1985. Mulbple-species rearing diets Pages 217 in Handbook of Insect Rearing, Volume I, Elsevier, Amsterdam.