The genetic diversity of cocoa has been studied using morphological, enzymatic and molecular descriptors. It has often proved difficult to obtain a clear classification due to factors including the significant genetic mixing that has occurred over the past three centuries, bias in the samples analysed (e.g. the limited number of Forastero samples collected in Peru for Witches’ broom resistance) the uncertain origin of some accessions (e.g. some early studies were based on material believed to be Criollo, but which was in fact of hybrid origin), different populations have been used in different studies. Breeding has been hampered by a lack of knowledge of the genetic diversity and level of heterozygosity of the accessions.
The main results of the diversity studies are:
Many breeding programmes have only used a limited number of Upper Amazon Forastero types collected by Pound. Genotypes from other populations have been used very little or not at aD (e.g. wild French Guiana, LCT EEN, Colombian EBC types, etc). It would be particularly interesting to set up prospective trials of crosses between genotypes from these different populations. This would exploit the diversity of natural T cacao populations that have not previously been used and may result in new heterotic combinations. Secondly, the genetic diversity studies have given useful information for population breeding approaches, such as reciprocal recurrent selection. Thirdly, the narrow genetic basis used in many cocoa breeding programmes to date is favourable for the exploitation of the expected linkage disequilibria within such populations. QTL mapping is generally done on a few specific progenies, and the results only relate to the clones involved. It is possible to enlarge such studies to analyse the degree to which genetic inkage between markers and traits of interest has been maintained during the evolution and domestication processes in genetic groups such as MC, SCA and MO, Forastero, Criollo, Trinitario or Nacional.
Introduction
A few reminders on the history of cocoa
According to Bartley (pers. comm.), the formation of hybrid populations following the introduction of Forastero types into original Criollo plantations may correspond to four main waves of introductions, possibly involving different genotypes. In addition to the introductions described above between Trinidad and Venezuela, there may also have been introductions of Ecuadorian cocoa into Mexico, introductions of varieties grown at the mouth of the Amazon into Central America, and of Amazon varieties into Colombian Criollo plantations in the 19th Century.
Selections were made in Trinidad among the Trinitario population and large numbers of clones (ICS, for Imperial College Selection) were distributed world-wide. The current plantations in Central America and Venezuela, which are very heterogeneous, often comprise a population of hybrids of varying degrees of introgression between Forastero and Criollo. The terms Criollo and Forastero originally came from Venezuela, where a distinction was made between traditionally grown local varieties (Criollo) and foreign trees (Forastero) introduced later from Trinidad, which were also known as Trinitario referring to the country from which they were introduced.
The Ecuadorian Nacional cocoa became an important variety in the second half of the 19th Century following the increase in cocoa consumption. According to Pound (1945), the “Nacional” type is probably indigenous to Eastern Ecuador. He refers to the existence of groups of very old wild cocoa trees resembling the Nacional type, known as “amacigales”, in primary forest areas cleared for new crops. Nacional cocoa differed from the wild cocoa trees found in the Amazon Valley. Some Nacional traits have more resemblance to Criollo than Forastero types, but Nacional also has characteristics that distinguish it from both groups. One such characteristic is that Nacional cocoa has the specific Arriba’ flavour (Enriquez 1993), Venezuelan cocoa types were introduced into Ecuador around 1890, via a few pods from Trinidad, where they had been introduced previously (Pound 1945). This material was particularly vigorous and precocious, even on poor soils, and most planters took seedlings from these trees to add to their original Nacional plantings. As a result, there was significant genetic mixing between the different origins, and pure types gradually disappeared (Soria 1970 a and b).
Cocoa was introduced into Africa more recently. It was first brought by Spanish or Portuguese seafarers, to Săo Tome in 1822 and Fernando Po in 1855 (Burle 1952). Swiss missionaries then made other introductions, from Suriname, and the first cocoa seeds were sown on the African mainland in 1857. The first material planted in West Africa, particularly Ghana, was Lower Amazon (Amelonado) type but this was followed by introductions of hybrid Trinitario and Criollo types from 1920 (Toxopeus 1972) which formed hybrids with original Amelonado type. However, each introduction comprised only a very limited number of genotypes, and the genetic basis of the cocoa populations initially grown in West Africa was very narrow. Moreover, the origin of the material is unclear.
Cocoa was introduced in the 16th century into Asia and the Pacific (\Wood 1991; Young 1994). In 1560, Venezuelan Criollo trees were introduced into Celebes by the Dutch, who later also introduced this type into Java. In addition, the Spanish introduced Criollo types from Mexico into the Philippines in 1614. In 1798, cocoa was taken by the British to Madras, India from the island of Amboina, and it was introduced into Ceylon (now Sri Lanka) from Trinidad at about the same time. From Ceylon, cocoa was subsequently transferred to Singapore and Fiji (1880), Samoa (1883), Queensland (1886), and Bombay and Zanzibar (1887). Cocoa was also grown in Malaysia as early as in 1778 and in Hawai by 1831.
Classification of cocoa
Morris (1882) was the first botanist to propose classification of cocoa populations into two groups: Criollo and Forastero. His classification was taken up by Pittier (1933), who designated each group as a different species: I lelocarpum for Forastero and I cacao for Criollo. However, all cocoa populations are inter-fertile, which in fact justifies the designation of a single species covering all wild and cultivated cocoa populations. Cuatrecasas (1964) proposed two sub-species: I cacao subsp. cacao for Criollo and I cacao subsp. sphaerocarpum for Forastero. A third group, Trinitario, contains hybrids between these two sub-species. This overall classification into two morpho-geographic groups, Criollo and Forastero, has been and is still in widespread use.
Classification of cocoa is difficult as it is affected by the history of cocoa domestication and by the substantial genetic mixing that has occurred, mainly over the last three centuries. The classifications proposed for the species I. cacao have therefore never been fully satisfactory. For example, according to some authors (Cheesman 1944; Soria 1970a and b) Nacional is an Upper Amazon Forastero, while for Enriquez (1993), some of its technological characteristics make it resemble a Criollo type.
Numerous hypotheses have also been put forward about the origin of the Criollo group (Cheesman 1944; Mora Urpi 1958; Cuatrecasas 1964; Purseglove 1968, Whitkus et a!. 1998). The most widely held are those of Cheesman and Cuatrecasas. Based on Vavilov’s principle, Cheesman (1944) considered the Upper Amazon as the centre of origin of Criollo and Forastero, given that it is in this region that the greatest morphological diversity is observed. Cheesman suggested that the spread of Criollo throughout Central America began from a small population in the upper reaches of the Amazon, which may have crossed the Andes with the help of man, and then formed differentiated populations as it spread. Cuatrecasas (1964), on the other hand, suggested that the species was indigenous from the Amazon region to Mexico.
Cuatrecasas backed up his hypotheses with observations of supposedly wild cocoa trees in the Lacandona forest near Chiapas, Mexico (Cuatrecasas 1964).
Study of the genetic structure of T. cacao
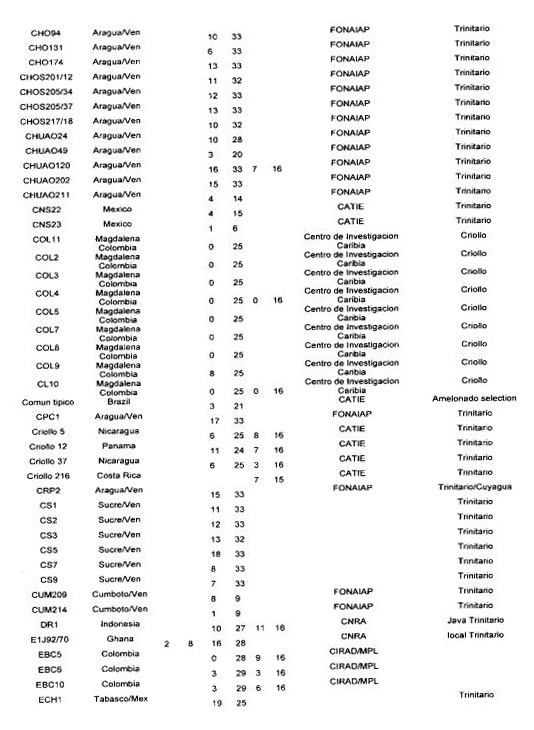
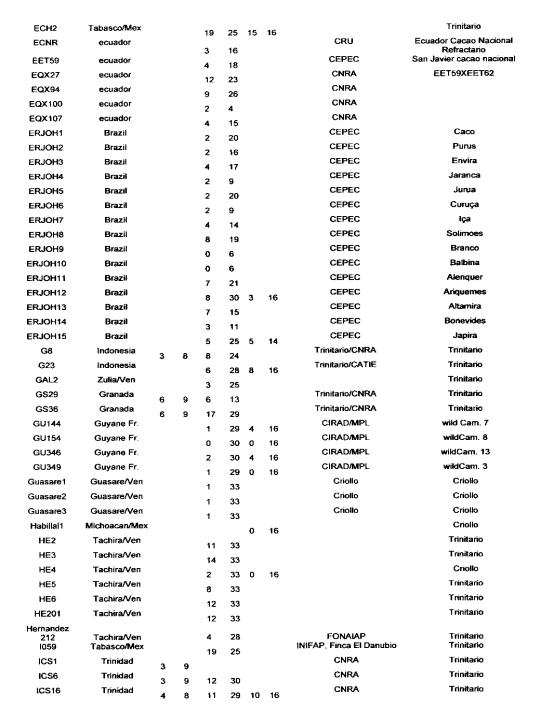
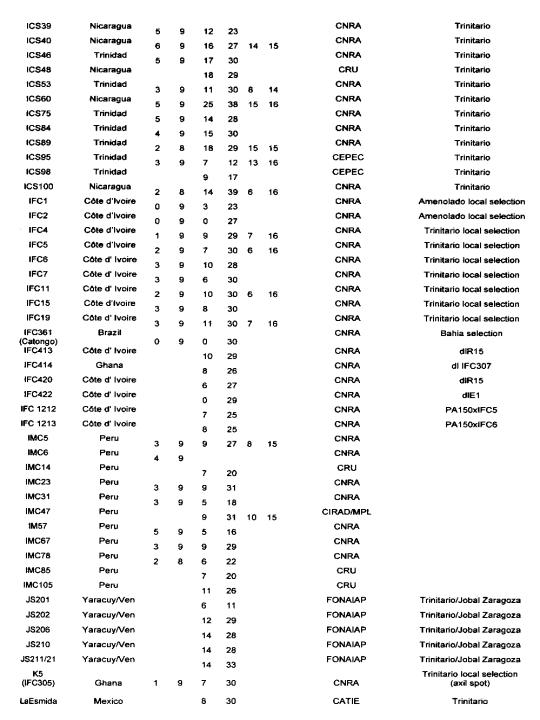
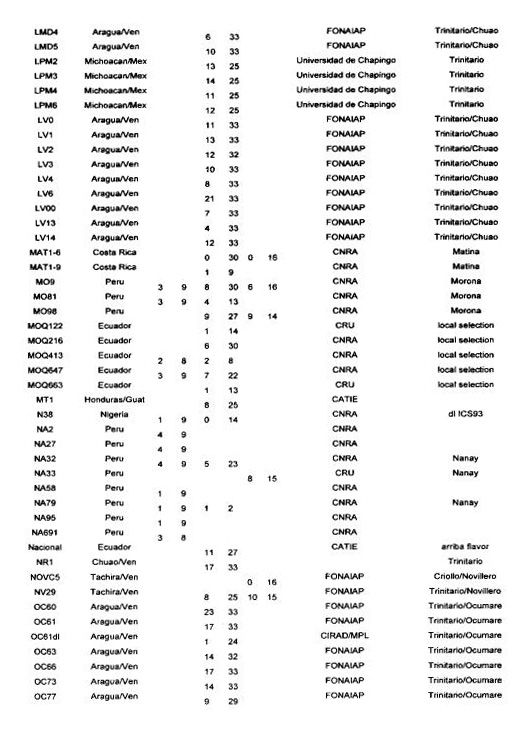
Many authors have participated in genetic diversity studies of T cacao with morphological, enzymatic or molecular markers (for a recent review see Lanaud of at 1999b). We report here some of the main results that have a direct bearing on cocoa genetic improvement. We firstly report on the level of heterozygosity as observed with isozymes, RFLP and microsatellite markers in the CIRAD laboratory at Montpellier.
Degree of heterozygosity of cocoa accessions
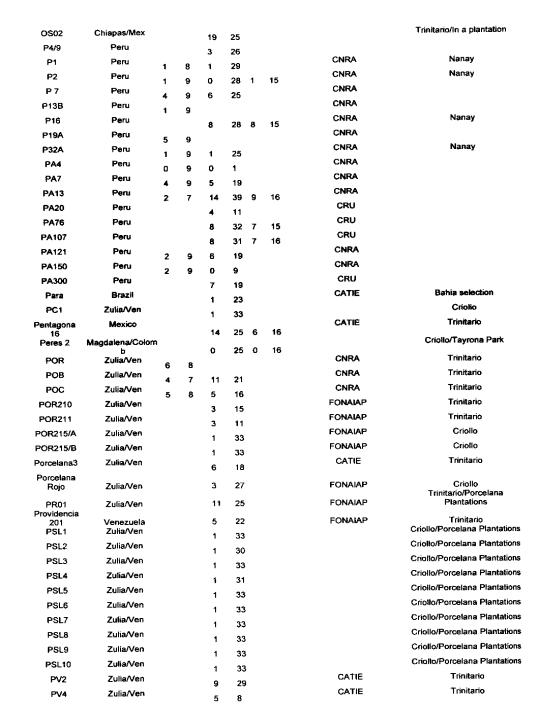
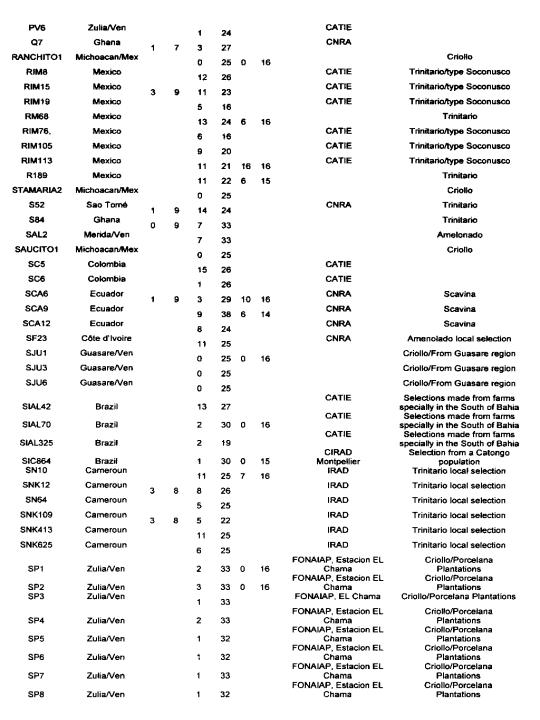
The data reported in Annex 1 are derived from the following sources: isozymes studies carried out by Lanaud (1987), RFLP studies carried out by Laurent et a!. (1994) and by Risterucci, Motamayor, Raboin and Lanaud (unpublished data), and microsatellite studies carried out by Motarnayor (also unpublished data).
The hybrid forms between Criollo and Forastero are called Trinitario” in Annex 1 even if they correspond to ~modem Criollo” varieties that correspond in fact to ancient Criollo more or less introgressed by Forastero genes as demonstrated by Motamayor et at (2000a and b).
The results show that most of Trinitar4o clones (e.g. UF, CS, UIT) are very heterozygous, which is in agreement with their hybrid origin. Some other populations displayed a higher degree of homozygosity, for instance wild Forastero from French Guiana (GU), Amelonado and Catongo Forastero (C361) varieties and certain Criollo clones (LAN, COL, Guasare, POR, PSL), corresponding to ancient cultivars. Certain Upper Amazon Forastero types also appeared to have a relatively high degree of homozygosity, for instance the EBC clones collected in Colombia, certain Ecuadorian LCTEEN clones and SCA 6.
These results are very valuable to cocoa breeders since they can be used to determine the clones most likely to create uniform hybrid progenies or to determine homozygous genotypes belonging to different genetic groups which could be crossed to exploit possible hybrid vigour.
The origin and diversity of Nacional, Criollo and Trinitario varieties
The analyses revealed a very small proportion of polymorphic Ioci among the individuals of ancient Criollo varieties. Moreover, within the group, hardly any molecular differences were observed, despite the highly contrasting morphotypes collected from Mexico to Venezuela, such as the Venezuelan Porcelana, Pentagona and Guasare, and the Criollo from the Lacandona Forest in Mexico.
The Criollo clones taken from collections generally appeared to be much more heterozygous. These clones had been originally selected not only for bean quality characters but also for agronornic characteristics (vigour, production or disease resistance). A Factorial Correspondence Analysis showed that the diversity of current
Criollo overlapped with that of the Trinitario clones studied. The more vigorous Criollo types may therefore correspond to ancestral Criollo forms into which Forastero genes have been introgressed to varying degrees.
Furthermore, the origin of the Forastero parents at the origin of modern Criollo and Trinitario was analysed. Around 90% of modern Criollo and Trinitario types apparently result from hybridisation and subsequent introgression between two genetically uniform types: homozygous Lower Amazon Forastero on one side and homozygous ancient Criollo on the other side. This is why in most cases only the same two alleles are found on each locus in the modern Criollo/Trinitario hybrid groups. Genotypes belonging to these groups would therefore represent different levels of recombinations of the Criollo and Lower Amazon Forastero parental genomes.
Ecuador/an Naciona!. Lerceteau et a!. (1997) used RFLP markers to study the Ecuadorian Nacional types which are grown today. A highly homozygous Nacional type was identified in old plantings in south-eastern and north-eastern Ecuador. These trees probably represent the homozygous ancestor at the origin of all the current hybrid varieties, (also called ‘Nacional’ types in collections), that result from Trinitario introgressions into this ancestral type with its extremely limited genetic base.
The diversity of Forastero populations. The Forastero group comprises a large number of wild populations and cultivated varieties originating from South America, which are found from Ecuador to the Guyanas. It includes vigorous trees and numerous sources of disease resistance. Significant genetic variability has been identified among these populations (e.g. Pound 1938 and 1945; Allen and Lass 1983), Certain Upper Amazon populations or geographic groups, such as LCTEEN and MC, seem to be the most variable (Sounigo et at 1996, see also this Proceedings), and there is continuous variation between them. Substantial within-population diversity has also been detected inside these populations, particularly those from Ecuador. Russel eta!. (1993) showed that within-population diversity was greater than between population diversity for three Peruvian and Ecuadorian populations (IMC, PA, LCTEEN). Using enzymatic markers, the GU types from French Guiana, on the other hand, do not seem to vary much and have a relatively high level of homozygosity (Lanaud 1987) although significant morphological diversity has been observed in terms of pod shape (Lachenaud and Sallee 1993).
However, it is difficult to compare the degree of diversity among these different populations. In effect, not all the populations were sampled or represented in the same way. The surveys made by Allen in Ecuador were not based on any strict criteria and covered a very large area in which material was taken from a large number of trees. Pound’s Peruvian clone samples, on the contrary, were taken for a precise purpose:
witches’ broom resistance. The material, which was taken from a limited number of trees is, therefore, not necessarily representative of the genetic diversity found in such area. Likewise, there was little material from clones from Colombia or from the Brazilian middle or upper Amazon in the material collected by Pound. Despite these often very biased samples, the genetic diversity (morphological, agronomical and by markers) observed was consistently substantial and greater than that found among the populations sampled in French Guiana or along the Orinoco in Venezuela.
The specificity of certain populations. Several populations or varieties seem to be clearly differentiated from the other morphogeographic groups identified for the species:
The narrow genetic base of the cocoa trees currently grown and used in improvement programmes. Genetic marker studies have shown the extremely narrow genetic basis of the Criollo, Trinitario, Nacional and Amelonado varieties, which account for almost all the traditional cocoa trees grown world-wide. From 1950 onwards, the Upper Amazon Forastero types collected by Pound started to be incorporated into cocoa improvement programmes. However, only a small part of the genetic diversity of the Pound collections has so far been used. In effect, it was somewhat by accident that some of them were distributed to a large number of producing countries (Lockwood and End 1993). Pods from nine of these clones, produced by Posnette in 1944 in an experiment on incompatibility designed for students, were sent to Ghana. The progenies were then spread throughout West Africa and later to Malaysia. Genetic improvement programmes have largely been, and continue to be, based on this Upper Amazon Forastero material. The material was generally hybridised with local Amelonado or Trinitario material, and later inter-Amazon crosses were also made. It is concluded therefore that only a small share of the genetic diversity of the species has been exploited, in terms of both traditional varieties and breeding programmes.
Consequences for cocoa genetic improvement
The first conclusion for cocoa breeding is that there is a large scope to increase the genetic bases for cocoa breeding. Certain Forastero types, such as GU clones, Ecuadorian LCTEEN clones and Colombian EBC clones have not yet been used for breeding purposes. Moreover, these genetic diversity analyses have also demonstrated the significant diversity of certain Upper Amazon populations, despite the fact that some samples were small or collected with precise criteria, such as the surveys made by Pound. This means that surveys of wild material should be continued in certain Upper Amazon regions such as Peru or Colombia, where there is substantial diversity that has not yet been sampled to any real extent.
Modern Criollo/Trinitario type varieties apparently result primarily from hybridization between two or three homozygous genotypes, and likewise, Nacional varieties apparently derive from a very small number of genotypes. This situation opens up a wide range of possibilities for improving these varieties, which produce an aromatic cocoa highly sought after for fine chocolate production. They could be improved for certain traits (like yielding capacity and precocity) by using existing or new hybrid combinations, with other genetic groups, selecting for favourable recombinants (e.g. combining good quality with good yield and disease resistance).
The possibilities for exploiting new types of hybrids are not limited to the improvement of Criollo or Nacional varieties, but are open to all cocoa breeding populations including hybrids between different Forastero populations. The hybrid vigour observed so far in many breeding programmes between Upper Amazon Forastero from Peru and Lower Amazon Forastero or Criollo could also exist, or be even more important, in many other hybrid combinations which have not yet been tested.
The new insights into the genetic structure of cocoa also have significant consequences for the choice and management of populations used in reciprocal recurrent selection strategies. In effect, sufficient genetic variability has to be maintained in the genetically different base populations to enable continuous genetic progress.
The narrow genetic base of the cocoa trees currently grown or being used for breeding purposes also favours exploitation of linkage disequilibria that may have been maintained between molecular markers and interesting traits during the genome evolution process. This phenomenon could enable greater use of the data acquired on the genome. For instance, in the case of the current Trinitario/Criollo, which have a very small number of parental genotypes, there have been very few generations of recombinations (probably six or seven at the most) between the first hybrids and current varieties. The close genetic linkages detected in some Trinitario clones between certain markers and genes of interest (Lanaud et al. 1999a; Lanaud et a!. 2000; Clement et a!. 2000; Paulin et a!. 2000) may have been maintained for most of this population. If this is the case, this will allow greater screening possibilities and exploitation of the Criollo/Trinitario group using the markers close to those genes.
As explained above, this situation could be similar for the new” Nacional varieties, i.e. the “pure” Nacional varieties which have received genes from certain Trinitario types. The small number of generations of recombinations has probably resulted in the maintenance of most of the close genetic linkages between markers and traits of interest, and should again enable screening of the varieties using those markers.
However, optimum use of the genetic resources of the species means clearly characterising all available accessions for morphological, agronomic, molecular, technological and sensorial traits. This comprehensive evaluation is highly complex but could be carried out firstly on a limited sample, as representative as possible of the diversity of the species using all available information (geographic, molecular and also morphological data).
One promising way for the future, with respect to a complete evaluation of cocoa genetic resources in terms of traits of interest, involves identification of the genes involved in the expression of those characters. This approach could be based on genome mapping, positional cloning and searching for candidate genes and would be associated with the use of high scale genome analyses. It would then be both easy and quick to look at the allele variations of the target gene within the species, identify original types and monitor the introgression of the character during breeding operations using a marker associated to the gene. This approach would thus facilitate the true exploitation of genetic diversity.
References
Allen J.B. and R.A. Lass. 1983. London Cocoa Trade Amazon Project. Final Report. Phase 1. Cocoa Growers’ Bufletin volume 34.
![]()

Biotech Glossary |
Bioinformatics |
Lab Protocol |
Notes |
Malaysia University |
Implications of New Insight into the Genetic Structure of Theobroma cacao L. for Breeding Strategies
Claire Lanaud, Juan-Curios Motan.ayor and Ange-Marie Risterucci
Cirad-Amis, TA 40/03, Avenue Agropolis, 34398 Montpellier Cedex 5, France
Cocoa was domesticated thousands of years ago by the Mayas and Aztecs (Paradis 1979). Even before the Spanish conquest, cocoa travelled along the trade routes used by the Mayas, Aztecs, and also the Pipil-Nicaraos (Young 1994; Coe and Coe 1996).
Criollo types then spread to Central America, and to a large number of Caribbean islands, including Trinidad in 1525 and thereafter to Jamaica. Cocoa introductions into Venezuela from Central America, particularly Costa Rica, were made by the Spanish (Pittier 1933), but it is also possible that cocoa may have been grown before the Spanish Conquest in the south-west of the country (Pittier 1933; Bergman 1969). The French planted cocoa in Martinique and Haiti, and the Portuguese planted it in Belem and Bahia around 1750, using Lower Amazon (Forastero) populations.
Hybridisations between Criollo and Forastero types in the 18th Century resulted in Trinitario types. It is not well known how this hybrid group initially formed. According to Pound (1945), the two populations could have met and hybridised on the islands of the Orinoco delta, including Trinidad and the Orinoco valley. This could have involved the cultivated Criollo population from Venezuela and the Amelonado type Forastero from Guyana, and was probably linked to exchanges between Venezuela and Trinidad. Cheesman (1944) reported that, in 1727, the ‘Blast”, perhaps a cyclone or an epidemic, destroyed the Criollo plantations in Trinidad. Plantations were reconstituted using seeds from the Orinoco valley, in Eastern Venezuela (Ciudad Bolivar). According to Cheesman, there is still some doubt as to the nature of these introductions, which may have been either Amelonado type Forastero, or already hybrid Trinitario forms. Other varieties, such as those grown on the Venezuelan coast, may also have been introduced (Batley, pers. comm.). Hybrids were produced by open pollination, and their superiority in agronomic terms and better resistance to diseases and pests favoured their use in Trinidad as a replacement for Criollo types. Trinitario material was distributed from Trinidad 70 years later, particularly into Venezuela.
The species Theobroma cacao (2n = 2x = 20) comprises a large number of highly morphologically variable populations, which can all be crossed with each other. Populations may be mostly autogamous or allogamous, depending on their genetic origin. A system of gametophytic-sporophytic self-incompatibility studied by several authors (Knight and Rogers 1955; Bouharmont 1960; Cope 1962; Glendinning 1962) increases the allogamy of certain populations.
Criollo and Trinitario varieties. The origin and diversity of Criollo and Trinitario have been studied by Motamayor et at (2000a and b). For this study, plant material was collected from the oldest plantings in Venezuela, irrespective of agronomic criteria, and in the Lacandona Forest, Mexico, near Maya archaeological sites where there are sub-spontaneous cocoa trees that probably descend from the cocoa trees grown by the Mayas. Samples were also taken in Yucatan. These representatives of pure Criollo varieties grown in the past have different pod shapes: oval with smooth surface, like those of Porcelana, or elongated with a very rough surface, like those of Pentagona. The analysis of this material was supplemented with that of so-called current’ Criollo and Trinitario varieties obtained from representative collections in Venezuela, Mexico and Costa Rica.
Bergmann J.F. 1969. The distribution of cacao cultivation in pre-Columbian America. Annals of the Association of American Geographers 59: 85-96.
Bouharmont J. 1960. Recherches cytologiques sur a fructification et l’incompatibilitę chez Theobroma cacao L. PubI. de l’lnearc, Serie Scientifique 59: 117 pp.
Burle L. 1952. La production du cacao en Afrique Occidentale Française. Centre de Recherches agronomiques de Bingerville. Bulletin 5: 3-21.
Cheesman E.E. 1944. Notes on the nomenclature, classification and possible relationships of cocoa population. Tropical Agriculture 21: 144-159.
Clement 0., AM. Risterucci, L. Grivet, J.C. Motamayor, J.A.K. N’Goran and C. Lanaud. 2000. Cartographie genetique de caractéres lies au rendement, a Ia vigueur, a Ia résistance a
P. palmivora et aux feves, chez Theobroma cacao L.. In 13me Conference Internationale sur Ia Recherche Cacaoyére, Kota Kinabalu, Malaisie, 9 au 14 October 2000. In press.
Coe S.D. and M.D. Coe. 1996. The true history of chocolate. Thames and Hudson Ltd. London, United Kingdom.
Cope F.W. 1962. The mechanism of pollen incompatibility in Theobroma cacao. Heredity 17: 157 pp.
Cuatrecasas J. 1964. Cacao and its allies: a taxonomic revision of the genus Theobroma. Contributions from the United States Herbarium 35: 379-614.
Enriquez GA. 1993. Characteristics of cacao “Nacional” of Ecuador. Pages 269-278 in Proceedings of the International Workshop on Conservation, Characterization and Utilisation of Cocoa Genetic Resources in the 21st Century. Port-of-Spain, Trinidad. 13-17th
September. The Cocoa Research Unit, The University of the West Indies.
Glendinning DR. 1962. Natural pollination of cocoa. Nature 193, no. 4822: 1305.
Knight R. and H.H. Rogers. 1955. Incompatibility in Theobroma cacao. Heredity 9:69-77.
Lachenaud P. and S. SaIls. 1993. Les cacaoyers spontanes de Guyane. Localisation, ecologie et morphologie. Café, Cacao, The, XXXVII, n~ 2: 101-114.
Lanaud C. 1987. Nouvelles donnees sur a biologie du cacaoyer (Theobroma cacao U: diversite des populations, systemes d’incompatibilite, haploTdes spontanés. Leurs consequences pour lamélioration genetique de cette espéce. Doctorat d’etat, Paris Xl.
Lanaud C., I. Kebe, AM. Risterucci, 0. Clement, J.A.K. N’Goran, L. Grivet, M. Tahi, C. Cilas,I. Pieretti, A.B. Eskes and D. Despreaux. 1999a. Mapping quantitative trait loci (QTL) for resistance to Phytophthora palmivora in T. cacao L. Pages 99-105 in Proceedings of the l2~ International Cocoa Research Conference, November 17-23, 1996. Salvador, Bahia, Brazil.
Lanaud C., J.C. Motamayor and 0. Sounigo. 199gb. Le cacaoyer. Pages 141-173 in Diversité genetique des plantes tropicales cultivees. (Hamon, P., Seguin, M., Perrier, X., Glaszmann, J.C., eds). Collection repéres/CIRAD, Montpellier, France.
Lanaud C., M.H. Flament, S. Nyasse, AM. Risterucci, 0. Fargeas, I. Kebe, 0. Sounigo, L. Motilal, J.M. Thevenin, D. Paulin, M. Ducamp, D. Clement, J.A.K. N’Goran and C. Cilas.
2000. Bilan de ‘etude des bases génetiques de Ia résistance du cacaoyer au Phytophthora a ‘aide de marqueurs moléculaires. ~ l3~ Conference Internationale sur Ia Recherche
Cacaoyere, Kota Kinabalu, Malaisie, 9-20 Octobre 2000. In press.
Laurent V., AM. Risterucci and C. Lanaud. 1993a. Chloroplast and mitochondrial DNA diversity in Theobroma cacao. Theor. AppI. Genet. 57: 61-88.
Laurent V., AM. Risterucci and C. Lanaud. 1993b. Variability for nuclear ribosomal genes within Theobroma cacao U. Heredity 71: 96-103.
Laurent V., AM. Risterucci and C. Lanaud. 1994. Genetic diversity in cocoa revealed by cONA probes. Theor. AppI. Genet. 68: 193-1 95.
Lerceteau E., T. Robert, V. Petiard and 0. Crouziuat. 1997. Evaluation of the extent of genetic variability among Theobroma cacao L. accessions using RAPO and RFLP markers. Theor. AppI. Genet. 95:10-19.
Lockwood C. and M. End. 1993. History, technique and future needs for cacao collection. Pages 1-14 in Proceedings of the Workshop on the Conservation, Characterization and Utilization of Cocoa Genetic Resources in the 21st Century, 13-17 September, 1992, Port-of- Spain. Trinidad. The Cocoa Research Unit, Port-of- Spain, Trinidad.
Mora Urpi J. 1956. Notas sobre el posible origen y Ia variabilidad del cacao cultivado en America Tropical. Turrialba 8: 34-43.
Morris D. 1682. Cocoa: how to grow and how to cure it. Jamaica pp 1-45.
Motamayor J.C., AM. Risterucci, A.P. Lopez and C. Lanaud. 2000a. Domestication du cacaoyer. I. Preuve d’une origine sud americaine des cacaoyers cultives par les Mayas. In 13eme Conference Internationale sur Ia Recherche Cacaoyere. Kota Kinabalu, Malaisie, 9-20 Octobre 2000. In press.
Motamayor J.C., AM. Risterucci, C. Lanaud. 200Gb. Domestication du cacaoyer. II.Germplasm a rorigine des cacaoyers Criollo/Trinitario cuftivés avant 1950. In l3~~Conference Internationale sur Ia Recherche Cacaoyere. Kota Kinabalu, Malaisie, 9-20 Octobre 2000. In press.
NGoran J.A.K., V. Laurent, AM. Risterucci and C. Lanaud. 1994. Comparative genetic diversity of Theobroma cacao L. using RFLP and RAPD markers. Heredity 73: 589-597.
Paradis L. 1979. Ue cacao précolombien: Monnais d’echange et breuvage des dieux, Journ. d’Agric. Trad. et de Bota. AppI. XXVI: 3-4.
Paulin D. and A.B. Eskes. 1995. Le cacaoyer: strategies de selection. Plantations Recherche Developpement 2: 5-18.
Paulin D., AM. Risterucci, M. Ducamp, J.A.K. N’goran and C. Uanaud, 2000. Identification de QTL pour Ia résistance du cacaoyer a Ia pourriture brune des cabosses (Phytophthora palmivora). ~fl l3~ Conference Internationale sur Ia Recherche Cacaoyere, Kota Kinabalu, Malaisie, 9 au 14 Octobre 2000. In press.
Pittier H. 1933. Degeneration of cacao through natural hybridization. Journal of Heredity 36: 385-390.
Pound F.J. 1938. Cacao and witchbroom disease (Marasmius perniciosus) of South America. Archives of Cocoa Research 1: 20-72.
Pound F.J. 1945. A note on the cocoa population of South America. Pages 131-1 33 in Report of the Cocoa Conference held at Grosvenor House, London.
Purseglove J.W. 1968. Theobroma cacao L.. Pages 571-599 in Tropical crops. Dicotyledons 2. (Purseglove, J. W. (ed)). John Wiley and Sons Inc. London, United Kingdom.
Russel JR., F. Hosein, R. Waugh and W. Powell. 1993. Genetic differentiation of cocoa (Theobroma cacao U.) populations revealed by RAPD analysis. Molecular Ecology 2: 89-97.
Soria J. 1970a. Principal varieties of cocoa cultivated in Tropical America. Cocoa Growers’ Bulletin 19:12-21.
Soria J. 1970b. The present status and perspectives for cocoa cultivars in Latin America. Proceedings of the Trop. Reg. Amer. Soc. For Hod. Sci. Vol. 14. Miami, Florida.
Sounigo 0., Y. Christopher and R. lJmaharan. 1997. Genetic diversity assessment of Theobroma cacao L. using isoenzymee and RAPD analyses. Pages 35-51 in Annual Report for the Cocoa Research Unit for 1996, CRU, Port of Spain, Trinidad and Tobago.
Toxopeus H. 1972. Cocoa breeding: a consequence of mating system, heterosis and population structure. Pages 3-12 in Proceedings of the Conference on Cocoa and Coconuts in Malaysia — Incorporated Society of Planters! Kuala Lumpur, Malaysia.
WhEtkus R., M. de Ia Cruz and L. Mota-Bravo. 1998. Genetic diversity and relationships of cocoa (Theobroma cacao L.) in southern Mextco. Theor. AppI. Genet. 96: 621-627.
Wood GAR. 1991. A histo~, of early cocoa introductions. Cocoa Growers’ Bulletin 44: 7-12.
Young AM. 1994. The chocolate tree: a natural history of cacao. Smithsonian Institution Press, Washington, UnEted States, 200 pp